КРОВЕТВОРЕНИЕ
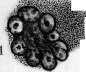
(В
i*^ ^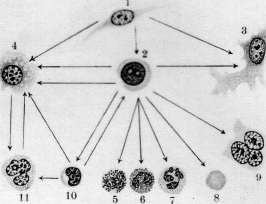

К ст. Кроветворение.

M*>i£**.fb.
ф| # ©р Чадит $ фМк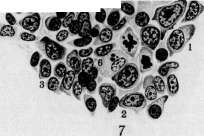
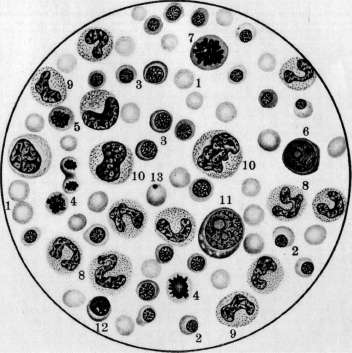
К ст. Кроветворение.
бластов, имеют более крупные размеры, чем эритроциты, а их протоплазма окрашивается смесью кислых и основных красок в промежуточный тон. Благодаря их крупным размерам их называют иногда макро-бластами. Эритробласты лежат в паренхиме костного мозга небольшими кучками клеток, находящихся в одинаковых стадиях созревания (рис. 6). Механизм проникновения эритроцитов из паренхимы костного мозга в просвет синусов не вполне выяснен.—Г етеропластический зрит* ропоэз, т. е. развитие проэритробластов из индиферентной.клетки, еще не вполне ясен. По взгляду унитаристов (Максимов), гемоцитобласт костного мозга может дифе-ренцироваться в различных направлениях, в том числе и в проэритробласта (рис. 5). При этом кариокинетическое деление является необходимым, т. к. во время него происходит внутренняя перестройка ядра клетки, к-рая становится способной развиваться лишь в эритроцита. По учению дуалистов (Негели) молодой эритробласт (пронормоб-ласт) развивается из индиферентной мезен-химной клетки, а Шридде производит его из эндотелия кровеносных сосудов. Эллер-ман (Ellermann) говорит об особи материнской клетки эритробластическ. ряда, о т. н. эритрогонии. Опыты эксплянтации лейке-мической крови доказывают, что гемоцитобласт способен в некоторых случаях in vitro диференцироваться в сторону эритро-бластов (Тимофеевский и Беневоленская).— Пат. увеличение эритропоэза наблюдается гл. обр. при анемиях (рис. 8). При этом содержание эритробластов в костном мозгу увеличивается, количество митозов возрастает, гетеропластический эритропоэз усиливается, жировой костный мозг замещается красным.. При некоторых особенно тяжелых анемиях, напр. злокачественном малокровии, развивается экстрамедуляр-ное К. (см. ниже гетеротопное кроветворение). При сильных интоксикациях и инфекциях может произойти угнетение эритропоэза: число эритробластов падает, фигуры деления их исчезают (арегенеративные— апластические анемии). Экспериментально подавление эритропоэза удается вызвать повторными кровопусканиями. Уменьшение эритропоэза наблюдается далее при лейкозах, особенно—острых. При злокачественном малокровии наблюдается развитие в костном мозгу наряду с нормобластами очень крупных гемоглобинсодержащих клеток—-мегалобластов и мегалоцитов. Эрлих(ЕпгИсп) высказал взгляд, что мегалобластическое превращение костного мозга является возвратом к эмбриональному К., а мегалобла-сты идентичны с первичными эритробласта-ми. Эта гипотеза нашла поддержку среди многих гематологов. Гетеропластическое развитие мегалобластов не может считаться вполне выясненным. По Негели, они развиваются из мезенхимных клеток, по Фер-рата (Ferrata) — из гемогистиобластов (гистиоцитов), по Ламбену (Lambin)—из ре-тикуло-эндотелия. Кроме того нарушение нормального эритропоэза может выражаться: 1) в распаде ядра эритробластов на отдельные сегменты; 2) в сохранении в эритро- бласте остатков ядерной субстанции (тельца Жолли, кольца Кабота); 3) в полихромазии и базофильной "точечности эритроцитов и наличии в них ретикуло-филаментозной субстанции, окрашиваемой основными красками в нефиксированных препаратах; 4) в недостаточной выработке НЬ. Некоторые из этих отклонений встречаются иногда в нормальной крови, но особенно сильно выражены при анемиях. Лейкопоэз (нормальный и патологический). 3 е р ни сты е лейкоциты развиваются в норме исключительно в костном мозгу преимущественно гомопластическим способом, т. е. путем размножения и созревания молодых зернистых клеток, так наз. миелоцитов (рис. 5, 6 и 8). Эти крупные клетки имеют круглое или бухтообразное ядро и зернистую протоплазму. Соответственно характеру зернистости различают нейтрофильные, эозинофильные и базофиль-ные миелоциты. Созревание миелоцитов в соответствующие сегментированные лейкоциты проходит через несколько стадиев, более подробно изученных для нейтрофи-лов. Ядро клетки делается сперва подковообразным (метамиелоцит Паппенгейма или юный нейтрофил, по Шиллингу), затем получает палочковидную форму в виде изогнутого, густо окрашивающегося жгута (палочкоядерный нейтрофил, по Шиллингу) и наконец распадается на несколько (2—5 и больше) сегментов, соединенных узкими перемычками (сегментированный нейтрофил). Начиная со стадия метамиелоцита, размножение клеток прекращается. Гетеропластический гранулопоэз, являющийся обычным при эмбриональном К., во взрослом организме в норме встречается редко. Считается доказанным, что все 3 вида миелоцитов происходят из общей родоначаль-ной клетки—гемоцитобласта (миелобласта). Другие, менее употребительные названия для этой клетки: лимфоидоцит(Паппенгейм), базофильный миелоцит (Dominici), лимфо-идная костномозговая клетка (Тюрк). Унитаристы (Максимов) помимо названия «гемоцитобласт» обозначают ее также большим лимфоцитом, тогда как дуалисты (Негели) придерживаются названия «миелобласт». Гемоцитобласт—клетка крупных размеров с круглым или слегка бухтообразным ядром, хроматин которого на сухих окрашенных мазках имеет вид тонкой равномерной сеточки с мелкими узловыми утолщениями. В ядре имеется несколько ядрышек. Протоплазма базофильна, незерниста, количество ее невелико. Гемоцитобласт дает положительную реакцию на протеолити-ческий фермент, оксидазу и пероксидазу— в этом видят отличие его от морфологически близко стоящего к нему большого лимфоцита или лимфоблаота, не дающего этих реакций. Во взрослом организме количество гемоцитобластов невелико, у новорожденных их значительно больше, а в эмбриональном периоде они преобладают над остальными клетками. Гемоцитобласт при созревании в миелоцита должен, по Максимову, подвергнуться митозу, во время которого и происходит диференцировка дочерних клеток в том или ином направлении (рис. 5). При этом базофилия протоплазмы уменьшается, количество последней увеличивается, в ней иногда развивается густая азурофильная зернистость, хроматиновые глыбки ядра делаются более грубыми, а ядрышки менее отчетливыми. Затем начинает вырабатываться тот или другой вид специфической зернистости, сперва в незначительном количестве, обычно около ядра на месте бухтообразного углубления. Такие переходные формы от гемоцитобластов к миелоцитам выделяются под названием про-миелоцитов. При дальнейшем созревании промиелоцита азурофильная зернистость, если она имелась, исчезает, базофилия протоплазмы постепенно ослабевает, специфическая зернистость увеличивается, причем у эозинофилов часто наряду с оксифильными зернами появляются отдельные базофиль-ные, неметахроматические зерна. Кроме того допускается, особенно при лейкозах, развитие миелоцитов непосредственно из индиферентной мезенхимной клетки (гемо-гистиобласт Феррата, рет.-энд. клетка) без промежуточного стадия гемоцитобласта. С другой стороны гемоцитобласт, подобно тому как у эмбриона, и во взрослом организме может развиваться из индиферентной мезенхимной клетки (см. ниже гетеротоп-ное кроветворение). Выход зрелых гранулоцитов в просвет синусоидов костного мозга объясняется их амебоидной подвижностью; здесь также может играть роль изменение давления крови в синусоидах, а следовательно и их просвета. Повышение интенсивности гранулопоэза наблюдается при многих инфекциях и особенно при миелозах. При нейтрофильном гиперлейкоцитозе усиливается выработка нейтрофилов и может наступить превращение жирового костного мозга в красный. В зависимости от силы и продолжительности вызывающей причины, клеточный состав костного мозга может резко меняться, и гетеропластиче-ский гранулопоэз значительно усиливается. Усиленное размножение эозинофильных миелоцитов и увеличенное образование их из гемоцитобластов наблюдаются при эози-нофильном гипер лейкоцитозе. При миелозах происходит превращение жирового костного мозга в миелоидный с развитием в паренхиме его более молодых гранулоцитов, а в острых случаях—гемоцитобластов. При хрон. инфекциях, интоксикациях и особенно при миелозах наряду с превращением жирового костного мозга в красный происходит развитие островков миелоидной ткани и в других местах организма (см. ниже гетеротопное кроветворение). Подавление гранулопоэза наблюдается при многих интоксикациях организма, при некоторых тяжелых инфекциях, при злокачественном малокровии. Костный мозг при этом беднеет гранулоцитами, и преобладающим элементом в нем может оказаться гемоцитобласт. Обычно это сопровождается дегенеративными изменениями гранулированных лейкоцитов и фокусами некроза паренхимы костного мозга. Отклонения от нормы в' развитии гранулоцитов могут состоять: 1) в недостаточной вы- работке специфической зернистости или даже полном отсутствии ее; 2) в неравномерном созревании ядра и протоплазмы; 3) в ненормальной дольчатости ядер гемоцитобластов (Ридеровские формы); 4) в развитии т. н. нейтрофильных близнецов, т. е. нейтрофилов, имеющих два ядра, совершенно-одинаково построенных. Особенно сильное отклонение от' нормы в гранулопоэзе наблюдается при острых миелозах: почти полное отсутствие гранулопоэза, причем преобладающим элементом в паренхиме костного мозга является гемоцитобласт. Иногда он имеет мелкие размеры, близко стоя по-своей морфологии к малому лимфоциту (ми-кромиелобласт).Из дегенеративных форм гранулоцитов заслуживают внимания дегенеративные формы палочкоядерных нейтрофилов, появляющихся в костном мозгу при так называемом дегенеративном ядерном сдвиге (Шиллинг). Образование лимфоцитов крови происходит в лимфоидной ткани, гл. обр. в лимф, узлах и селезенке, преимущественно-гомопластическим путем (рис. 7). Обычно считается, что малые лимфоциты крови происходят путем деления больших лимфоцитов, к-рые поэтому часто называются лимфобластами (Негели), реже—лим-фогониями Benda. Впрочем, по Максимову, гораздо чаще наблюдается размножение* средних—мезо-лимфоцитов. Деление лимфоцитов происходит в лимф, узлах, гл. образ.. в центре размножения фоликулов (рис. 7). Различают две фазы фоликулов лимфат. узла: деятельную и покойную. В течение первой—фоликул крупного размера содержит много больших и средних лимфоцитов' с многочисленными митозами; в течение второй—клеточных элементов в нем мало, преобладают малые лимфоциты. Г е т е р о-пластическое образование лимфоцитов происходит из индиферентных клеток мезенхимы. При этом фиксированная клетка мезенхимы делится митотически, давая происхождение круглым клеткам, имеющим светлые ядра и слабо базофильную протоплазму. При дальнейшем размножении последних получаются типичные средние лимфоциты. Этот процесс особенно выражен при переходе фоликула лимф, узла из фазы покоя в фазу деятельности (Максимов) . При развитии новых лимфатических фоликулов лимфоциты образуются из таких же индиферентных клеток мезенхимы. Это образование происходит преимущественно на месте бывших жировых долек. Генетические отношения фиксированных и свободных клеток лимфоидной ткани, по Максимову, можно схематически выразить следующим образом: ^ Средние и ->- Малые ^--' большие "*~ лимфо-Недиференцированный ' лимфоциты циты синцитий лимфоидной ~~-~^ ткани """-"-». Фиксиро- -> макро- ванные ги- фаги стиоциты При пат. условиях Максимов допускает кроме того возможность развития из больших и средних лимфоцитов элементов миелоидной ткани, из малых лимфоцитов—ми- кромиелоцитов, а из тех и других—моноцитов и макрофагов. Возможность происхождения моноцитов из свободных гистиоцитов (макрофагов) также допускается Максимовым. Патол. лимфопоэз, наблюдаемый при некоторых инфекциях, интоксикациях и особенно лимфаденозах, может состоять в довышенном образовании больших лимфоцитов—лимфобластов, у взрослого в крови нормально не встречающихся, далее—в ненормальной дольчатости ядер больших лимфоцитов (Ридеровские формы), в развитии лимфоцитов, почти лишенных протоплазмы или, наоборот, с обильным количеством последней, в отсутствии азурофильной зернистости, нормально встречающейся у одной трети лимфоцитов. Усиленный лимфопоэз сопровождается гиперплазией лимф, ткани с увеличением лимфатических узлов, селезенки, фоликулов слизистых оболочек, зобной .железы и с развитием новых лимфом в различных местах организма. Угнетение лим-•фопоэза происходит вследствие разрушения лимфойдной ткани лимфат. узлов; при этом кровь беднеет лимфоцитами (лимфо-гранулема, лимфосаркоматоз). Происхождение моноцитов крови не вполне выяснено. Имеется три главных взгляда: 1) моноциты происходят из рети-куло-эндотелиальн. аппарата путем мобилизации рет.-энд. клеток (Ашоф, Кийоно, Шиллинг); 2) моноциты развиваются в миелоид-ной системе из миелобластов (Негели); 3) моноциты образуются из лимфоцитов путем гипертрофии ядра и протоплазмы последних и представляют переходную форму от лимфоцита к макрофагу или полибласту (Максимов, Bloom). Такая гипертрофия лимфоцита происходит в кровеносных сосудах с замедленным током крови. Впрочем Максимов не отрицает происхождения моноцитов из ретикуло-эндотелия. Моноцитоз крови обычно сопровождается гиперплазией ретикуло-эндотелиального аппарата и мобилизацией ретикуло-эндотели-альных клеток, которые могут появляться в крови, особенно при некоторых хронических сепсисах. Эти клетки носят все признаки гистиоцитов или макрофагов и дают все возможные переходы к обычным моноцитам. Все это говорит за роль ретикуло-эндотелия в происхождении моноцитов; вместе с тем и лимфоцит, как показывают опыты эксплантации, может в самое короткое время перейти в моноцит. Тромбоцитопоэз (нормальный и патологический). У низших позвоночных и у птиц тромбоциты являются настоящими клетками, снабженными ядрами. У млекопитающих и человека кровяные пластинки представляют собой безъядерные образования, происходящие согласно исследованию Райта (Wright) из отделившихся частичек протоплазмы мегакариоцитов костного мозга. Тромбоцитопоэз происходит ловидимому таким образом, что мегакариоцит посылает в просвет синусоидов костного мозга амебоидные отростки азурофильной протоплазмы; от них отрываются мелкие частицы, к-рые и уносятся током крови в виде кровяных пластинок (рис. 1) (см. Виццоцеро бляшки). Впрочем нек-рые авторы до наст. времени придерживаются старых взглядов о происхождении кровяных пластинок, производя их то из протоплазмы лейкоцитов и эритроцитов то из ядер лейкоцитов.— Кровяная пластинка, по Шиллингу, представляет собой измененное ядро эритро-бласта, выталкиваемое из молодого эритроцита уже в кровяном русле. Повышенное развитие мегакариоцитов из гемоцитоблас-тов и усиленное функционирование их сопровождается повышением числа кровяных пластинок в крови и появлением гигантских форм (хлороз, миелозы). Иногда в кровь попадают части мегакариоцитов с обломками ядра. Некоторые патол. процессы сопровождаются тромбоцитопенией и появлением патологических пластинок с ненормальной зернистостью или отсутствием последней или с сильно выраженной базо-филией краевой зоны или пластинок ненормальных размеров (злокачественное малокровие, апластические анемии, «эссенциаль-ная тромбопения»). . Отношения между кроветворными органами и кровью. Морфолог, состав крови зависит от двух факторов: от производства форменных элементов и от их потребления, причем оба эти процесса находятся между собой в теснейшей взаимной связи и взаимно друг друга обусловливают. Повышение эритропоэза находит отражение в составе крови появлением регенеративных форм эритроцитов и эритробластов. Но при сильных и быстрых раздражениях костного мозга появление эритробластов в крови может быть вызвано простым вымыванием их из миелои-дной ткани вследствие нарушения кровообращения в ней. При мегалобластозе костного мозга в крови появляются мегалоци-ты и мегалобласты. Отсутствие в крови регенеративных форм эритроцитов при наличии тяжелой анемии позволяет предполагать отсутствие эритропоэза в костном мозгу (см. также Анемия). Отношения, существующие между лейкопоэзом и составом лейкоцитов крови, являются более сложными, так как распределение лейкоцитов в сосудистом ложе и процессы потребления лейкоцитов могут протекать независимо для каждого вида. При быстро развивающемся нейтрофиль-ном лейкоцитозе подвижные нейтрофиль-ные лейкоциты могут в самое короткое время эмигрировать из паренхимы костного мозга в кровеносные сосуды. При хрон. воспалительных процессах без осложнений, протекающих при почти полном отсутствии ядерного сдвига, костный мозг богат зрелыми и юными нейтрофилами, а при тяжелых острых инфекциях с сильным ядерным сдвигом он оказывается состоящим гл. обр. из промиелоцитов (Шиллинг). При лейкозах явления гиперплазии кроветворных органов в большинстве случаев находят отражение в лейкемич. составе крови, и по морфологии последней можно судить о морфологич. составе кроветворных органов, и наоборот. В.общем однако следует сказать, что состав крови далеко не всегда отражает процессы, протекающие в кроветворных органах; существует какой-то механизм, пока еще не выясненный, который, смотря по обстоятель- ствам, облегчает или затрудняет переход форменных элементов из костного мозга в периферическую кровь (костномозговой барьер). (См. также Лейкоцитозы, Лейкемия, Лейкоцитарная формула.) Гетеротопное К. При пат. условиях (ин-. фекции, интоксикации, тяжелые малокровия, миелозы) наблюдается возникновение миелоидной ткани в разных местах организма, т. н. экстрамедулярный миелопоэз. Миелоидная метаплазия как правило возникает сначала в селезенке, затем в печени, лимф, узлах, надпочечниках и других местах организма. При этом вначале появляются ми елоциты, за.теммегакариоциты, позднее и не всегда—эритробласты. Экстраме-дулярное происхождение миелоидных элементов в наст, время большинством объясняется соответствующей диференцировкой местных элементов (аутохтонная теория). Колонизационная теория, по к-рой миелоидная метаплазия возникает из клеток, заносимых током крови в различные места организма, в настоящее время большинством отвергается. Вопрос о том, из каких местных клеток развиваются элементы миелоидной ткани, является одним из наиболее спорных в гематологии. Часть авторов (Шридде, Герцог, Негели) производит их из клеток сосудистой стенки. Близко к этому стоит учение Феррата, Гоффа (Hoff), Сысоева и друг., приписывающее гемопоэ-тическую способность ретикуло-эндотелию. По Феррата, элементы миелоидной ткани развиваются из ретикуло-эндотелия, пройдя стадии гемогистиобласта и гемоцито-бласта. Меллендорф (Mollendorff) считает, что зернистые лейкоциты могут развиваться из фибробластов; это мнение впрочем встречает возражения большинства ученых. Доминичи, Доуни, Вайденрейх (Downey, Weidenreich), Максимов наделяют лимфоцитов гемопоэтическими способностями. По Максимову, гистиоциты и сосудистый эндотелий не обладают способностью к гемопоэзу, и экстрамедулярный миелопоэз происходит или за счет лимфоцитов крови и тканей или же из индиферентных мезенхимных клеток. Теории кроветворения. Вопрос о генетических отношениях форменных элементов крови друг к другу и клеточным формам соединительной ткани является и до наст. времени одним из самых спорных в гематологии. Здесь можно различать четыре главнейших направления, или четыре теории кроветворения: 1) унитарное направление. 2) умеренно-унитарное, 3) дуалистическое и 4) триалистическое. Унитарное учение (Гравиц, Максимов, Вайденрейх, Хлопин, Мясоедов) полагает, что все незернистые элементы крови и кроветворных органов, т. е. малые лимфоциты, большие лимфоциты, гемоцитобласты, а также (по нек-рым) и моноциты, являются недиференцированными клетками, способными при известных условиях давать происхождение зернистым лейкоцитам, эритроцитам и мегакариоцитам. Согласно этому учению гемоцитобласт (миелобласт) и большой лимфоцит способны давать происхождение одинаковым клеткам, а существую- I щие между ними биохим. и морфол. различия непостоянны и зависят исключительно от временных условий существования. Унитарное учение основывается главн. обр. на экспериментальных исследованиях, особенно—Максимова и его учеников. Согласно этому учению в раннем эмбриональном периоде все клетки мезенхимы обладают полнотой гемопоэтических способностей, во взрослом же организме эта способность сохраняется за отдельными недиференцированными клетками мезенхимы, к к-рым, по нек-рым, относится также и ретикуло-эндо-телий. По Максимову, часть клеток соединительной ткани односторонне диференциро-вана (фибробласты, эндотелий кровеносных и лимф, сосудов), часть (клетки ретикуло-эндотелиального аппарата и вообще гистиоциты) сохранила только некоторые способности (способность развиваться в фибробласты и в моноциты крови) и наконец часть сохранила все эмбриональные свойства с широкими гемопоэтическими возможностями (рисунок 2). Учение умеренных унитаристов (Паппенгейм) производит все элементы крови из общего родоначальника—лимфоидоцита (гемоцитобласта). Лимфоциты крови и лимфатической-ткани являются уже диферен-цированными элементами, неспособными к гемопоэзу. По учению дуалистов (Эрлих, Негели, Шридде) лимфоциты и зернистые лейкоциты имеют двух родоначальников—лимфобласта и миелобласта, к-рые ни при каких условиях не могут переходить друг в друга и различаются по морфологическим и биохимическим особенностям. Связующим звеном для элементов крови, по Негели, является индиферентная мезенхимная клетка, которая обладает всей полнотой гемопоэтических потенций. По триалистическому учению гистиоциты, как свободные, так и фиксированные, составляют 3-й независимый кроветворный орган, подобный миелоидной и лимф, системе организма. Этот орган производит моноциты крови, которые так. обр. по своему происхождению являются независимыми от лимфоцитов и зернистых лейкоцитов (Aschoff). Культура нормальной и пат. крови. Культура лейкоцитов крови in vitro, впервые произведенная Авроровым и Тимофеевским,. дала возможность выяснить нек-рые спорные вопросы гематологии. Зернистые лейкоциты нормальной крови как клетки с законченным развитием в течение нескольких дней подвергаются in vitro распаду, тогда как лимфоциты и моноциты развиваются в короткое время в фагоцитирующие поли-бласты или макрофаги. Этим подтверждается учение Максимова о роли незернистых лейкоцитов в происхождении клеточных: форм в воспаленной ткани. В более старых культурах макрофаги вытягиваются и постепенно переходят в типичные фиброциты, вырабатывающие тоно-фибрилы и колаге-новые волокна, — развивается настоящая соединительная ткань (Максимов). При своем развитии в полибласта лимфоцит крови I проходит через стадий моноцита, что гово- 54а рит за возможный генезис моноцитов из лимфоцитов (Максимов, Блюм, Тимофеев-ский и Беневоленская). Проявление гемо-поэтической способности со стороны лимфоцитов и моноцитов нормальной крови in vitro не происходит. В этом отношении эти клетки отличаются от гематоцитобласта (миелобласта) лейкемической крови: гемо-цитобласты в самое короткое время дифе-ренцируются in vitro в гранулоциты, реже—в эритробласты [см. отдельную таблицу (ст. 531—532), рисунки 3 и 4, и отдельную таблицу (ст. 659—660), рисунок 15] (Тимофеев-ский и Беневоленская). Сопоставление указанных двух факторов говорит не в пользу унитарного учения. Лит.—см. лит. к ст. Гематология и Костный мозг. А. Тдмофеевский. Смотрите также:
Смотрите также:
- КРОВОИЗЛИЯНИЕ, скопление крови, излившейся из сосудов, в полостях организма или в тканях. Полостные К. обозначают соответственно названию полости, содержащей излившуюся кровь (см. Haemothorax, Гемоперикард, Haematometra, Haematosal-pinx, Haematocele, Haematomyelia и пр.). ...
- КРОВООБРАЩЕНИЕ. Содержание: I. Физиология. План построения системы К....... 543 Движущие силы К............ 545 Движение крови в сосудах........ 546 Скорость ...
- КРОВООСТАНАВЛИВАЮЩИЕ СРЕДСТВА, haemostatica, haemostyptica, весьма большая группа разнообразных веществ, действие которых остается невыясненным до наст. времени. К. с. подразделяются на 1) местные или наружные, действующие лишь при нанесении их непосредственно на ...
- КРОВОПУСКАНИЕ, искусственное быстрое удаление из кровеносной системы значительных количеств крови при помощи венесекции, венепункции, артериотомии или пункции артерий. С терап. целью обычно производится К. в 200—500 см3 крови. Однако нек-рые ...
- КРОВОСМЕШЕНИЕ, инцест, термин для обозначения половой связи между,близкими родственниками, запрещаемой законом. В прежнее время очень много говорили о вреде К. для потомства; самые тяжелые б-ни (калеки, идиоты, глухонемые; душевнобольные, слепые) ...